Article Text

Abstract
Objectives To seek evidence for the role of hypoxia in early human tendinopathy, and thereafter to explore mechanisms whereby tissue hypoxia may regulate apoptosis, inflammatory mediator expression and matrix regulation in human tenocytes.
Methods Fifteen torn supraspinatus tendon (established pathology) and matched intact subscapularis tendon (representing ‘early pathology’) biopsies were collected from patients undergoing arthroscopic shoulder surgery. Control samples of the subscapularis tendon were collected from 10 patients undergoing arthroscopic stabilisation surgery. Markers of hypoxia were quantified by immunohistochemical methods. Human tendon-derived primary cells were derived from hamstring tendon tissue obtained during hamstring tendon anterior cruciate ligament reconstruction. The impact of hypoxia upon tenocyte biology ex vivo was measured using quantitative real-time PCR, multiplex cytokine assays, apoptotic proteomic profiling, immunohistochemistry and annexin V fluorescence-activated cell sorter staining.
Results Increased expression of hypoxia-inducible factor 1α, Bcl-2 and clusterin was detected in subscapularis tendon samples compared with both matched torn samples and non-matched control samples (p<0.01). Hypoxic tenocytes exhibited increased production of proinflammatory cytokines (p<0.001), altered matrix regulation (p<0.01) with increased production of collagen type III operating through a mitogen-activated protein kinase-dependent pathway. Finally, hypoxia increased the expression of several mediators of apoptosis and thereby promoted tenocyte apoptosis.
Conclusion Hypoxia promotes the expression of proinflammatory cytokines, key apoptotic mediators and drives matrix component synthesis towards a collagen type III profile by human tenocytes. The authors propose hypoxic cell injury as a critical pathophysiological mechanism in early tendinopathy offering novel therapeutic opportunities in the management of tendon disorders.
Statistics from Altmetric.com
Overuse tendon injuries, namely tendinopathies, pose a significant, highly prevalent problem in musculoskeletal medicine.1 The intrinsic pathogenetic mechanisms underlying the development of tendinopathies are largely unknown; however, excessive apoptosis (programmed cell death) and inflammation have recently been shown to be functionally important in several model systems.2,–,4 The interaction between apoptosis and inflammation is critical to tissue homeostasis in a variety of tissues and is known to play a key role in inflammatory articular,5 neoplastic,6 neurodegenerative7 and cardiovascular diseases.8
Apoptosis may be induced by many stimuli, including hypoxia. The magnitude and duration of the hypoxic microenvironment largely determines whether cells become apoptotic or adapt and survive.9 Hypoxia-inducible factor 1α (HIF-1α), is a critical regulator in this process. Whereas HIF-1α can initiate apoptosis by inducing high concentrations of proapoptotic proteins,10 such as caspase 3, during hypoxia, HIF-1α induces anti-apoptotic proteins,11 such as clusterin and Bcl-2, while the proapoptotic protein Bax is downregulated. HIF-1α is also recognised to control key aspects of inflammation, particularly leucocyte recruitment and subsequent cytokine production.12
Biopsy and in-vivo model studies suggest that tendon hypoperfusion13 and subsequent hypoxia14 are central elements of tendon failure, but few data have addressed this at a mechanistic level in humans. In rodent models, cyclic strain regulates the expression of the HIF-1α in rat tendon fibroblasts.15 Increased hypoxia in the tenosynovium of rheumatoid arthritis patients is observed compared with controls,16 and Benson et al17 recently described hypoxic damage throughout the spectrum of rotator cuff disease. To date, no studies have explored the functional role of hypoxia in modulating apoptosis, inflammation and matrix regulation in tendinopathy.
One of the major limitations of human studies is that tendon biopsies are usually obtained when patients are symptomatic and therefore biopsy material probably represents chronic, rather than early phase, disease. Medical intervention at this early stage may offer considerable therapeutic advantage over later surgical approaches. We previously demonstrated that matched subscapularis tendons from patients with full thickness rotator cuff tears represent a model of early human tendinopathy based on histological appearances and significantly increased levels of cytokines and apoptotic markers in these tissues.18 The purpose of this study was to assess formally the degree of hypoxic markers within this early phase human model ex vivo, and thereafter to explore the mechanistic outcome of tissue hypoxia on apoptosis, inflammation and matrix production in tenocytes in vitro.
Methods
Human model of tendinopathy
All procedures and protocols were approved by the ethics committee under ACEC no. 99/101. Fifteen supraspinatus tendon samples were collected from patients with rotator cuff tears undergoing shoulder surgery (table 1). The mean age of the rotator cuff ruptured patients was 55 years (range 38–70 years)—the mean tear size was 2.3 cm2. Samples of the subscapularis tendon were also collected from the same patients. Patients were only included if there was no clinically detectable evidence of subscapularis tendinopathy on a preoperative MRI scan or macroscopic damage to the subscapularis tendon at the time of arthroscopy—by these criteria they represented a truly preclinical cohort. An independent control group was obtained comprising 10 samples of subscapularis tendon collected from patients undergoing arthroscopic surgery for shoulder stabilisation without rotator cuff tears. The absence of rotator cuff tears was confirmed by arthroscopic examination. The mean age of the control group was 32 years (range 17–38 years).
Patient demographics and rotator cuff tear size
Tissue collection and preparation
Arthroscopic repair of the rotator cuff was carried out using the standard three-portal technique as described previously.19 The cross-sectional size of the rotator cuff tear was estimated and recorded as described previously.20 The subscapularis tendon was harvested arthroscopically from the superior border of the tendon 1 cm lateral to the glenoid labrum. The supraspinatus tendon was harvested from within 1.5 cm of the edge of the tear before surgical repair without surgical debridement. For immunohistochemical staining the tissue samples were immediately fixed in 10% (v/v) formalin for 4–6 h and then embedded in paraffin. Sections were cut to 5 cm thickness using a Leica-LM microtome (Leica Microsystems, Wetzlar, Germany)and placed onto Superfrost ultra plus glass slides (Gerhard Menzel, Braunschweig, Germany).The paraffin was removed from the tissue sections with xylene, rehydrated in graded alcohol and used for histological and immunohistochemical staining as per previously established methodologies.
Human tendon-derived cells were explanted from hamstring tendon tissue of five patients (aged 18–30 years) undergoing hamstring tendon anterior cruciate ligament reconstruction. All of these samples showed normal fibrotendinous tissue architecture and no evidence of tendinopathy. Cultures were maintained at 37°C in a humidified atmosphere of 5% carbon dioxide for 28 days. Cells were subcultured and trypsinised at subconfluency, cells from the third and fourth passage were used under either normoxic or hypoxic conditions.
Hypoxic conditions
The disaggregated cells were filtered through nylon mesh, and were plated at 1×106/ml into 75 cm2 culture flasks (BD Falcon, Leuven, Belgium) under normoxic (21% oxygen) or hypoxic (1% oxygen) conditions using an air-tight hypoxic incubator with inflow and outflow valves (Wolf Laboratories Ltd, York, UK). Oxygen concentrations were continuously measured with a built-in oxygen sensor and the percentage of oxygen was adjusted by the addition of nitrogen.
Histology and immunohistochemistry techniques
Sections were stained with H&E and toluidine blue for determination of the degree of tendinopathy as assessed by a modified version of the Bonar21 score (grade 4 = marked tendinopathy; grade 3 = advanced tendinopathy; grade 2 = moderate degeneration; grade 1 = mild degeneration; grade 0 = normal tendon). This included the presence or absence of oedema and degeneration together with the degree of fibroblast cellularity and chondroid metaplasia. Thereafter, sections were stained with primary monoclonal antibodies directed against the following markers: HIF-1α, clusterin, Bcl-2 (R&D Systems Europe, Oxfordshire, UK), BNIP-3 and vascular endothelial growth factor (Abcam, Cambridge, UK) CD68 (tissue macrophages), CD3 (T cells), CD4 (T helper cells), CD206 (M2 macrophages) and mast cell tryptase (mast cells).
Endogenous peroxidase activity was quenched with 3% (v/v) hydrogen peroxide, and non-specific antibody binding blocked with 2.5% horse serum in TBST buffer for 30 min. Antigen retrieval was performed in 0.01M citrate buffer for 20 min in a microwave. Sections were incubated with primary antibody in 2.5% (w/v) horse serum/human serum/TBST at 4°C overnight. After two washes, slides were incubated with the Vector ImmPRESS reagent kit (Vector Laboratories, Peterbourough, UK) as per the manufacturer's instructions for 30 min. The slides were washed and incubated with Vector ImmPACT DAB chromagen solution for 2 min, followed by extensive washing. Finally, the sections were counterstained with haematoxylin. Positive (human tonsil tissue) and negative control specimens were included, in addition to the surgical specimens for each individual antibody staining technique. The omission of primary antibody and use of negative control isotypes confirmed the specificity of staining.
We applied a scoring system based on previous methods21 to quantify the immunohistochemical staining. Ten random high power fields (×400) were evaluated by three independent assessors (NLM, JHR, ALC) blinded to diagnosis/tissue origin. In each field the number of positive and negatively stained cells were counted and the percentage of positive cells calculated giving the following semiquantitative grading: grade 0 = no staining; grade 1 = less than 10% of cells stained positive; grade 2 = 10–20% of cells stained positive; grade 3 = more than 20% of cells positive.
Apoptosis and matrix regulation
Normoxic or hypoxic cells were analysed using annexin V binding, which detects the transfer of phosphatidylserine to the outside of the membrane, cells were harvested with trypsin and stained with the annexin V–FITC apoptosis detection kit (BD Pharmagen, San Diego, California, USA) for flow cytometric analysis. Cells were also assessed using the Tunel assay (R&D Systems Europe) for DNA fragmentation as per the manufacturer's instructions. The production of 35 apoptotic-related proteins in normoxic or hypoxic cell culture supernatants was assessed using a proteome profiler (R&D Systems Europe), which contains capture and control antibodies spotted in duplicate on nitrocellulose membranes. The array is incubated with a cocktail of biotinylated detection antibodies.
Normoxic or hypoxic tenocytes were evaluated for immunocytochemical staining of collagen I, collagen III and MMP13 (Abcam) to assess tenocyte matrix production. Total soluble collagen was measured from cell culture supernatants using the Sircol assay kit (Biocolor Ltd, Carrickfergus, Northern Ireland) according to the manufacturer's protocol. One millilitre of Sircol dye reagent was added to 100 µl test sample and mixed for 30 min at room temperature. The collagen–dye complex was precipitated by centrifugation at 10 000g for 10 min; and then washed twice with 500 µl ethanol. The pellet was dissolved in 500 µl alkali reagent. The absorbance was measured at 540 nm by a microplate reader. The calibration curve was set up on the basis of the collagen standard provided by the manufacturer. In addition, 10 samples further samples of torn supraspinatus, matched subscapularis and control biopsy were used to assess collagen protein quantification (ηg/ml) using collagen I and III human ELISA kits (Biorbyt, Carrickfergus, UK)as per the manufacturer's instructions.
Signalling experiments
The phosphorylation status of mitogen-activated protein kinases (MAPK), extracellular signal regulated kinases (ERK1/2), c-Jun N-terminal kinases (JNK) and p38 isoforms were evaluated using the human phospho-MAPK array (R&D Systems Europe) as per the manufacturer's instructions. The p38 MAPK inhibitor (SB203580) and ERK inhibitor (FR180204) were purchased from CalbioChem (Merck KGaA, Darmstadt, Germany) and used at IC50 10 µM, a concentration previously determined to offer optimal specific inhibition relative to off-target effects.22
RNA extraction and quantitative PCR
The cells isolated from the normoxic and hypoxic experiments were placed in Trizol before messenger RNA extraction. QIAgen mini columns (Qiagen Ltd, Crawley, UK) were used for the RNA clean-up with an incorporated column DNAse step as per the manufacturer's instructions. Complementary DNA was prepared from RNA samples according to AffinityScript (Agilent Technologies, Santa Clara, California, USA) multiple temperature cDNA synthesis kit as per the manufacturer's instructions. SYBR green real-time PCR was performed using SYBR green mastermix (Applied Biosystems, Carlsbad, CA, USA). Before setting up the SYBR green the cDNA was diluted 1 in 5 using RNase-free water. Each sample was analysed in triplicate. Primers (Integrated DNA Technologies, Leuven, Belgium) were as follows: GAPDH, 5′–TCG ACA GTC AGC CGC ATC TTC TTT–3′ (F) and 5′–ACC AAA TCC GTT GAC TCC GAC CTT–3′ (R), COL1A, 5′–CAA TGC TGC CCT TTC TGC TCC TTT–3′ (F) and 5′–CAC TTG GGT GTT TGA GCA TTG CCT–3′ (R), COL3A, 5′–TAT CGA ACA CGC AAG GCT GTG AGA–3′ (F) and 5′–GGC CAA CGT CCA CAC CAA ATT CTT–3′ (R), MMP13, 5′–AAG GAC CCT GGA GCA CTC ATG TTT–3′ (F) and 5′–TGG CAT CAA GGG ATA AGG AAG GGT–3′ (R), HIF-1α, 5′–TTG GCA GCA ACG ACA CAG AAA CTG–3′ (F) and 5′–TTG AGT GCA GGG TCA GCA CTA CTT–3′ (R), HSP70, 5′–tgc tgg aca agt gtc aag agg tca (f) and 5′–TCC TCT TGT GCT CAA ACT CGT CCT–3′ (R), CASPASE 3, 5′–TCA TTA TTC AGG CCT GCC GTG GTA (F) and 5′–TGG ATG AAC CAG GAG CCA TCC TTT–3′, CASPASE 7, 5′–TTC CTC TTC GCC TAT TCC ACG GTT (F) and 5′–ATT CAC CCT GGT GAG GAT CTG CAT–3′ (R), BCL-2, 5′–TTT CTC ATG GCT GTC CTT CAG GGT–3′ (F) and 5′–AGG TCT GGC TTC ATA CCA CAG GTT–3′ (R), CLUSTERIN, 5′–AGG TGG TCG TGA AGC TCT TTG ACT–3′ (F) and 5′–TTT CTC CGC CAC GGT CTC CAT AAA–3′ (R).
Cytokine production
A 25-Plex human cytokine assay evaluated the quantitative determination of 25 separate human cytokines using Luminex (Luminex Corp, Austin, Texas, USA) technology. Supernatants (n=5, in triplicate) were removed from normoxic, hypoxic or treated cells and analysed for cytokine production.
Statistical analysis
Results are reported as mean values±SD. Analysis of variance followed by Tukey's test or Student's t test was applied to in-vitro studies. Analysis between individual ex-vivo groups was examined by analysis of variance followed by the Student–Newman–Keuls test or Student's t test using Sigma Stat, version 3.1. A value of p<0.05 was considered to be significant.
Results
Early tendinopathy is associated with increased hypoxic and apoptotic markers
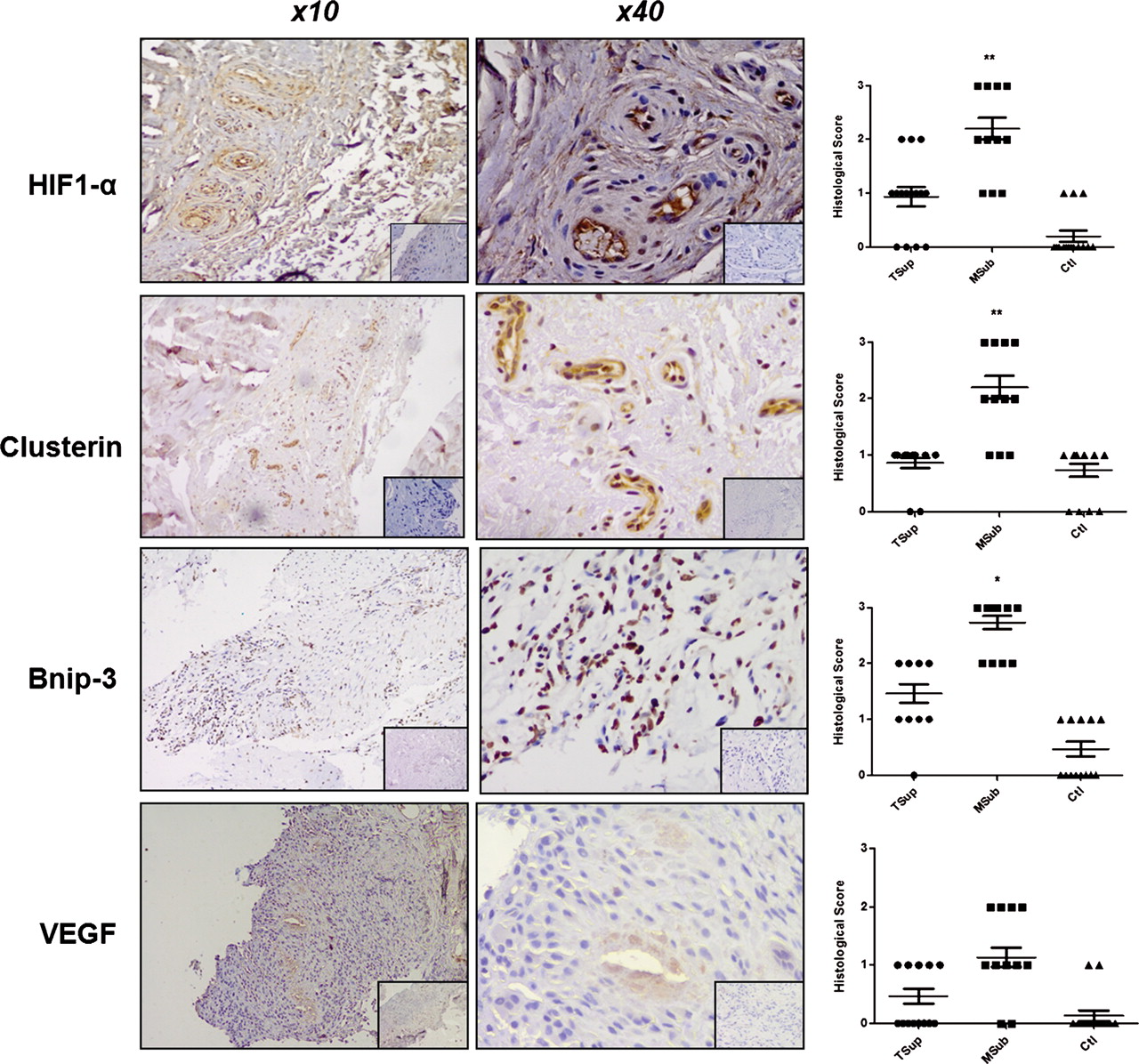
Torn supraspinatus samples exhibited marked degeneration, mucoid change and frank chondroid metaplasia (grade 4), whereas matched subscapularis tendon biopsies had grade 2–3 changes indicative of early tendinopathy. All control samples were classified as grade 1 consistent with normal fibrotendinous tissue with large distinct collagen fibrils. There were no significant correlations between the Bonar score and the mean duration of symptoms or patients' age. However, subscapularis tendon samples exhibited significantly greater staining for HIF-1α, clusterin and Bcl-2 compared with either matched torn supraspinatus samples or control tissue (figure 1). Subscapularis tendon samples exhibited significantly greater (p<0.01) numbers of macrophages, mast cells and T cells compared with either matched torn supraspinatus samples and control tissue. Inflammatory cell infiltrate and apoptosis markers correlated inversely (r=0.4, p<0.01) to rotator cuff tear size in the torn supraspinatus biopsies, with larger tears showing a marked reduction in inflammatory cell types and hypoxic/apoptotic markers.

Apoptosis and hypoxic markers in tendon samples. Subscapularis tendon stained for hypoxia-inducible factor 1α (HIF-1α), clusterin, Bnip-3 and vascular endothelial growth factor (VEGF) (magnification ×10 and ×40). Isotype IgG in bottom right corner. Graphs illustrate relative expression of corresponding proteins in human tendon samples. Histological scoring system: grade 0 = no staining; grade 1 = less than 10% of cells positive; grade 2 = 10–20% of cells positive; grade 3 = more than 20% of cells positive. Data are displayed as mean ±SEM, n=15 for supraspinatus and matched subscapularis, n=10 for control group (*p<0.05, **p<0.01).
Hypoxic induces apoptosis in tenocytes in vitro
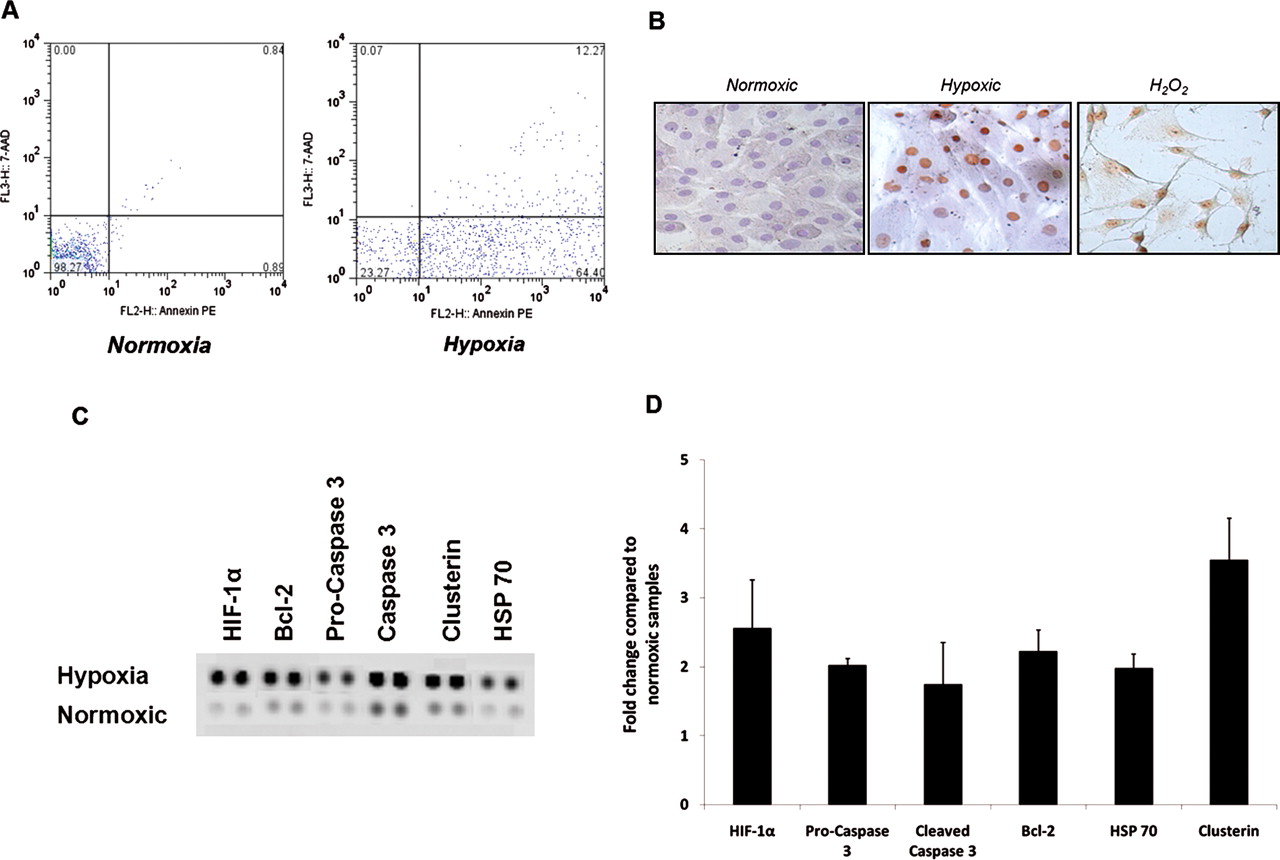
To explore the physiological significance of these findings, we first established an ex-vivo primary human tenocyte culture system in which ambient oxygen concentrations could be meticulously controlled as required. Hypoxia significantly increased the number of cells undergoing apoptosis assessed by annexin V fluorescence-activated cell sorter staining at 12 h (normoxia, mean 5±2% apoptosis; hypoxia, mean 23±8%), 24 h (normoxia, mean 3±2%; hypoxia, mean 58±10%) and 48 h (normoxia, mean 8±4%; hypoxia, mean 60±15%) (figure 2A; p<0.01). This was confirmed by Tunel staining showing increased levels of apoptosis in 1% conditions. Limited proteomic profiling revealed relative overexpression of Bcl-2, HIF-1α, clusterin, pro and cleaved caspase 3 and heat shock protein (HSP) 70, in hypoxic conditions (figure 2B; p<0.01). These protein expression changes were confirmed at the mRNA level by quantitative PCR that confirmed significantly elevated expression of HIF-1α, caspases 3 and 7, clusterin, Bcl-2 and HSP70 compared with normoxic conditions (figure 2C,D).

Apoptotic gene expression in hypoxic conditions. (A) Cell death after exposure to hypoxic conditions. Apoptosis was evaluated after treating tenocytes with 24 h of normoxic (24% oxygen) and hypoxic (1% oxygen) conditions. Flow cytometry profile represents annexin V–FITC staining in x axis and 7-AAD (Aminoactinomycin D) in y axis. (B) Cultured tendon fibroblasts underwent a Tunel assay to detect nuclear DNA degradation, a feature of apoptotic cells. Cells with brown-stained nuclei are positive for DNA degradation. Samples displayed include 24% oxygen, 1% oxygen and positive controls treated with hydrogen peroxide. Whole-cell lysates from normoxic (24% oxygen) and hypoxic (1% oxygen) conditions were harvested and apoptotic markers evaluated using an apoptotic proteome profiler (C). The fold change of apoptotic proteins was determined by densitometry and normalised to the control sample on the array (D). Data are shown as the mean fold change±SD of duplicate samples and are representative of experiments using five individual donors of tendon explant tissue. HIF-1α, hypoxia-inducible factor 1α; H2O2, hydrogen peroxide HSP70, heat shock protein 70.
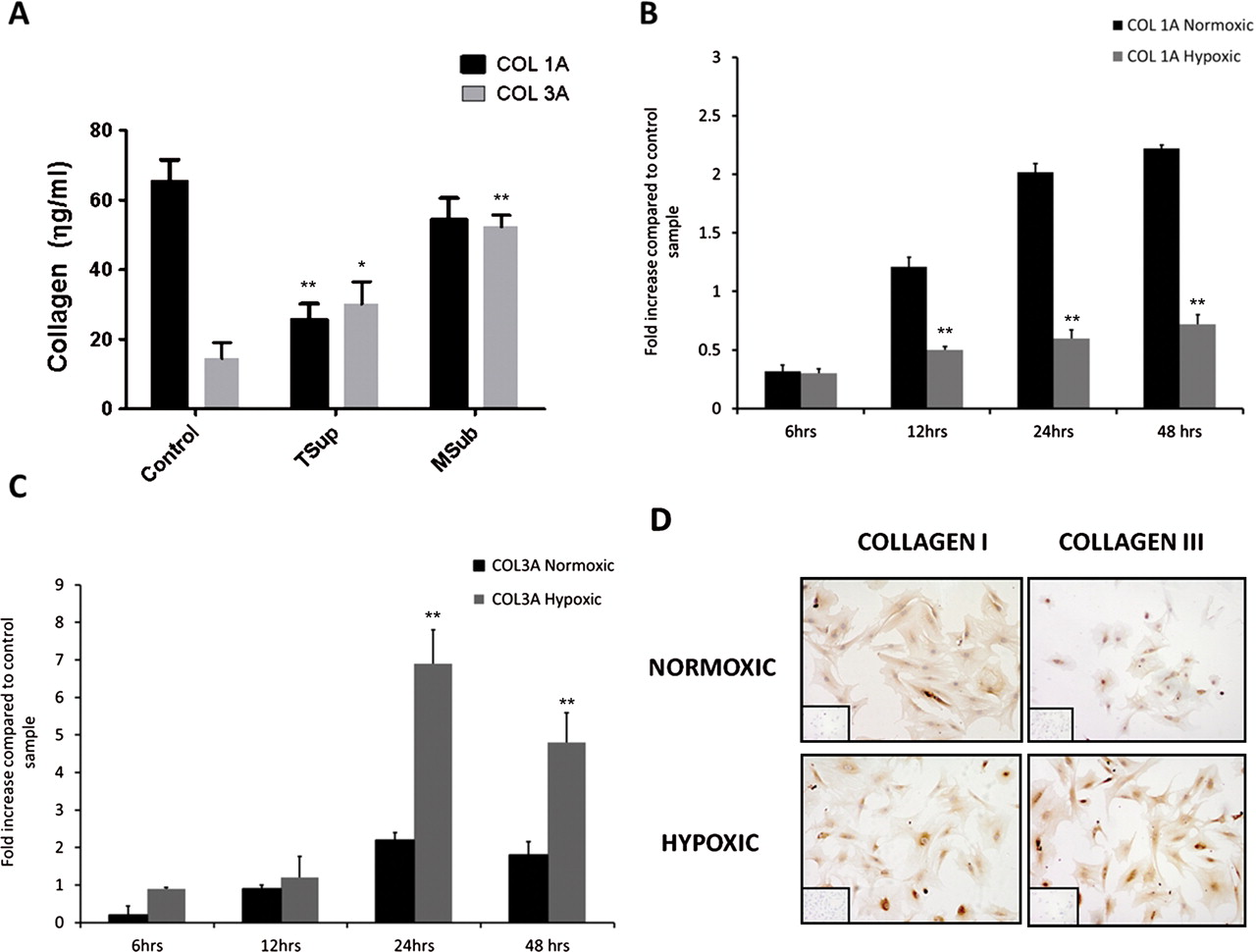
Hypoxia alters tenocyte collagen matrix regulation
As disproportionate type III collagen expression is a characteristic feature of tendinopathy, we next considered whether the hypoxic changes noted above might alter differential collagen synthesis by tenocytes and whether collagen matrix changes were present in tendon biopsies noted to have increased hypoxic markers. Collagen III was significantly (p<0.05) elevated in torn supraspinatus and matched subscapularis samples compared with controls (figure 3A). Collagen IA mRNA was significantly reduced at all time points in hypoxic compared with normoxic conditions. In contrast, collagen IIIA mRNA was significantly upregulated at 24 and 48 h in hypoxic compared with normoxic cultures (figure 3B,C). Commensurate with this at the protein level, hypoxia significantly elevated total collagen production compared with normoxia after 24 h (mean 328 µg/ml ±81 hypoxia; 520 µg/ml ±98 normoxic) and 48 h (712 µg/ml ±66 hypoxia; 909 µg/ml ±74 normoxic). Semiquantitative immunocytochemistry revealed no marked change in type I collagen protein expression in hypoxic compared with normoxic cultures, whereas collagen III staining was markedly increased at all time points by hypoxia (figure 3D).

Collagen production in vitro. (A) Collagen I and III concentrations (ηg/ml) were assessed using ELISA. Data represent mean collagen production (ηg/ml)±SD in 10 biopsies of torn supraspinatus, matched subscapularis and control tendon. The levels of mRNA for collagen type IA (B) and collagen type IIIA (C) were determined by real-time PCR. Data are shown as the mean±SD of triplicate samples and represent experiments on five individual patient samples. *p<0.05, **p<0.01 compared eith normoxic samples. (D) Immunostaining for collagen types I and III in normoxic and hypoxic conditions at 24 h incubation. Cells were plated onto chamber slides and incubated in different oxygen conditions for 24 h then fixed in methanol and stained with antibodies.
Hypoxia promotes cytokine production in tenocytes
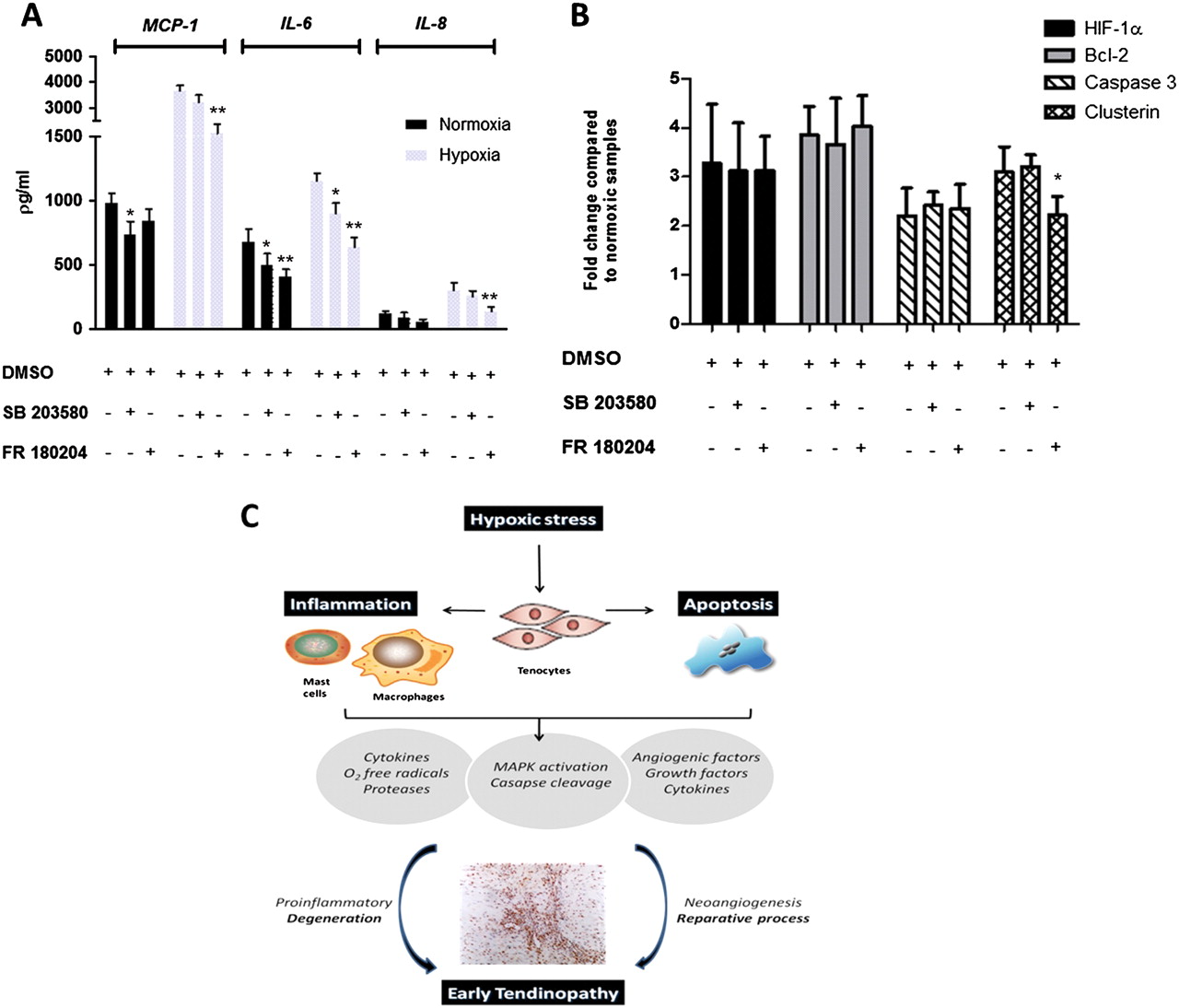
As our initial histological examination revealed a significant association between inflammatory infiltrate and hypoxic markers, we next explored the extent to which hypoxia could regulate the local cytokine milieu directly by modulating tenocyte behaviour. Hypoxic conditions rapidly induced significantly elevated production of interleukin (IL) 6, and of the chemokines IL-8 and MCP-1 commensurate with the notion that hypoxic tenocytes could initiate leucocyte recruitment (figure 4). In contrast, we found no production over time of a range of other cytokines including IL-4, IL-5, IL-10, IL-12, IL-13, IL-15, TNFα, IL-1β by tenocytes, suggesting that the earlier detection of such entities in early tendinopathy probably reflects infiltrating leucocyte populations.18

Effects of hypoxia on cytokine production in cultured tenocytes. Cultured tenocytes were incubated in a normoxic and hypoxic environment over a time course from 0 to 48 h. Data show levels of MCP-1, IL-8 and IL-6 in supernatants removed from culture at various time points analysed using Luminex. Data are shown as the mean±SD of triplicate samples and are representative of five individual experiments. *p<0.05, **p<0.01 compared with normoxic samples.
Hypoxia, MAPK expression and collagen synthesis
As expected, tenocytes cultured in a hypoxic environment exhibited a generalised increase in MAPK phosphorylation. In particular, we observed increased phosphorylation of ERK1, ERK2 and the α and β isoforms of p38 (figures 5A,B). To explore the functional consequences of this, we employed ERK and p38 MAPK inhibitors in our tenocyte cultures. We observed partial but significant abrogation of the hypoxia-induced increase in total collagen production by tenocytes (24 h dimethyl sulphoxide 592±32 µg collagen (mean±SD); p38 inhibitor 461±35 µg; ERK inhibitor 390±38 µg) (figure 5C). In contrast, p38 or ERK inhibition in normoxic conditions resulted in an increase in total collagen production (24 h dimethyl sulphoxide 402±33 µg collagen (mean±SD); p38 inhibitor 541±35 µg; ERK inhibitor 509±55 µg).
Collagen subtype synthesis was evaluated by real-time PCR. Both ERK and p38 inhibition abrogated the hypoxia-induced enhancement of collagen type III in tenocytes and significantly increased the relative expression of collagen III in normoxic conditions (figure 5D).
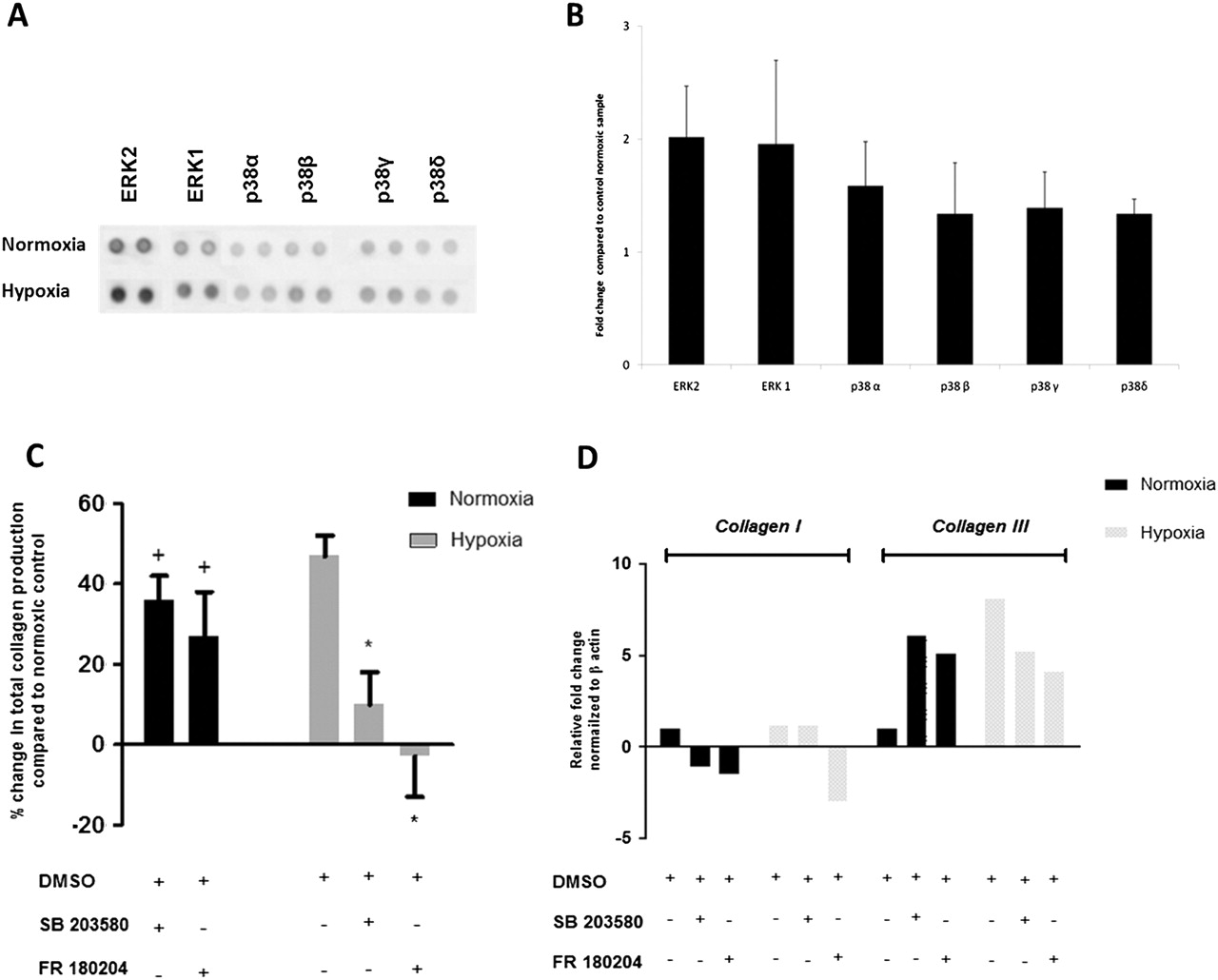
Hypoxia-induced phosphorylation of mitogen-activated protein kinase (MAPK) in cultured tenocytes and matrix response to p38 and extracellular signal regulated kinase (ERK) inhibition. Whole-cell lysates were examined for the phosphorylation of ERK1, ERK2 and p38 isoforms at 1 h after exposure to normoxic and hypoxic conditions (A). The fold change of MAPK was determined by densitometry and normalised to the control sample on the array (B). Data are shown as the mean fold change±SD of duplicate samples and are representative of experiments using five individual donors of tendon explant tissue. Cells were preincubated for 24 h with specific inhibitors for p38 (SB 203580), or ERK (FR 180204), which also included assay media. Total collagen production was assessed using Sircol assay (C). Data represent percentage change in total collagen production compared with normoxic control sample±SD in normoxic and hypoxic conditions over a 48 h time course. *p<0.05, **p<0.01 compared with normoxic samples. The levels of mRNA (D) for collagen type IA and collagen type IIIA were determined by real-time PCR. DMSO, dimethyl sulphoxide.
ERK and p38 inhibitors reduce proinflammatory cytokine/chemokine production but do not modulate apoptosis in tenocytes
Tenocyte production of IL-6, MCP-1 and IL-8 under normoxic and particularly hypoxic conditions was reduced by both p38 and ERK inhibition (figure 6A). ERK appeared more important in this regard. Annexin V staining revealed a moderate reduction (p=0.06) in the percentage of apoptotic cells in hypoxic conditions using either p38 (hypoxia control 62±10%, inhibitor 55±10%) and ERK inhibitors (hypoxia control 63±8%, inhibitor 52±12%). While ERK inhibition did induce a small but significant reduction in clusterin gene expression in hypoxic conditions, neither p38 and ERK inhibition had a significant effect on other key apoptotic genes in normoxic or hypoxic environments, suggesting that p38/ERK pathways do not significantly contribute to tenocyte apoptosis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The effects of extracellular signal regulated kinase (ERK) and p38 mitogen-activated protein kinase (MAPK) inhibition on cytokine production and apoptosis. (A) Cells were preincubated for 24 h in a normoxic and hypoxic environment with specific inhibitors for p38 (SB 203580), or ERK (FR 180204), which also included assay media. Data show levels of MCP-1, IL-8 and IL-6 in supernatants removed from culture analysed using Luminex. Data are shown as the mean±SD of triplicate samples and are representative of five individual experiments. (B) Whole-cell lysates were examined for the expression of hypoxia-inducible factor 1α (HIF-1α), caspase 3, Bcl-2 and clusterin, at 24 h after exposure to normoxic and hypoxic conditions with specific inhibitors for p38 (SB 203580), or ERK (FR 180204). Expression was determined by densitometry and normalised to the control sample on the array. Data are shown as the mean fold change±SD of duplicate samples and are representative of experiments using five individual donors of tendon explant tissue. (C) Schematic diagram illustrating the manner in which early tendinopathy may arise due to hypoxic injury. An increase in hypoxic stress that a tendon cell experiences results in the release of various inflammatory mediators with associated MAPK activation that interact to drive the tendon matrix toward a degenerative or reparative process. DMSO, dimethyl sulphoxide; O2, oxygen.
Discussion
Our study provides convincing evidence that hypoxia could operate as an initiator and thereafter a regulator of early tendinopathy. Herein we demonstrate that hypoxia-related proteins are present in early tendinopathy biopsies, and thereafter in mechanistic studies demonstrate that hypoxia regulates inflammatory and apoptotic mediators in tendon cells associated with a significant shift in collagen matrix synthesis.
Approximately 95% of collagen in normal tendon is type I; type III is present in small amounts under physiological conditions.23 Biopsies from normal and ruptured Achilles tendons demonstrate that ruptured tendons contain reduced quantities of type I collagen together with a significantly increased proportion of type III collagen,24 which in turn accounts for the decreased resistance of tendon to tensile forces and thereby subsequent rupture.25 Hypoxic cell damage has long been considered a mechanism underpinning degenerative tendon disorders.26 27 Conventionally, stress responses such as oxidative damage are considered to operate through MAPK pathways.28 These are upregulated by cyclical strain29 and are subsequently involved in apoptosis30 in tendinopathy; however, there remains limited investigation into signalling events involved in tendon biology. Our work suggests a functional role for ERK and p38 in the matrix and inflammatory response to hypoxia. Hypoxia not only increases the total collagen production but seems to ‘switch’ the production towards an increase in collagen III compared with type I, suggesting a detrimental phenotypical change in the extracellular matrix. In particular, ERK inhibition reduces the total collagen and collagen III production in hypoxia while it has the opposite effect (increased collagen and collagen III) in normoxia, suggesting that the ERK pathway is oxygen sensitive.
Experimental models provide good evidence for an early inflammatory response in tendinopathy.4 A running rodent model of tendinopathy is associated with the upregulation of key inflammatory modulators including the 5-lipoxygenase activating protein and cyclo-oxygenase,3 while rabbit and equine models show that excessive mechanical load induces acute inflammatory cell infiltrates.31 32 We have recently shown a distinct inflammatory infiltrate in early human tendinopathy33 and increased levels of proinflammatory cytokine in torn supraspinatus samples.18 Hypoxia has been shown to induce the expression of cytokines and proinflammatory molecules including platelet-derived growth factor, IL-6, IL-8 and platelet-activating factor in several animal and human models.34 35 Our data now demonstrate in human tenocytes that a hypoxic environment is capable of inducing key inflammatory cytokines that may ultimately disturb the balance between reparative and degenerative changes in the extracellular matrix. In particular, increased levels of MCP-1, IL-6 and IL-8, which have previously been shown to affect matrix metalloproteinase production in fibroblasts,36 probably have downstream effects on tenocyte collagen matrix production. Inhibition of ERK in hypoxia significantly reduced cytokine production and thus may provide a mechanism for reducing proinflammatory molecular events in tendinopathy. We and others have highlighted the key role of a large mast cell infiltrate in tendinopathy,33 37 38 and this combined with evidence from Gulliksson et al39 demonstrating hypoxic-induced IL-6 production in mast cells, adds growing importance to the role of chemokines in the initiation and perpetuation of the inflammatory cascade in tendon biology (figure 6C).
Dysregulated apoptosis has also been postulated as a primary cause of tendinopathies.40 Several causal factors, including mechanical overuse,41 hypoxia42 and oxidative stress43 have been proposed. We have previously shown increased expression of pro-apoptotic genes in a rodent overuse models and in torn human supraspinatus tendons compared with controls.44 Increased mechanical loading45 and loss of homeostatic tension41 increases tenocyte apoptosis in rodents and HIF-1α is induced during cyclical strain of tenocytes.13 In addition, the position of the rotator cuff tendons lends themselves to impingement and increased mechanical stresses, which are important anatomical factors in reduced oxygen tension.17 Increased HIF-1α has been shown in rotator cuff tears, although as these represent advanced disease by definition it is difficult to evaluate its relative role in early disease.17 Our study now reveals HIF-1α expression in early tendinopathy biopsies. Clusterin46 and Bcl-247 have been functionally implicated in DNA repair, cell cycle regulation, while both play a prosurvival role during cell death.48 Their abundant expression in early tendinopathy biopsies and overexpression under hypoxic conditions at both the protein and mRNA level strongly supports the concept of hypoxia as a regulator of apoptosis. Moreover, parallel changes were noted in collagen subtype synthesis in our studies. The equilibrium between pro/anti-apoptotic markers during hypoxic conditions in a tissue rescue/wound healing process may thus play a critical role in determining the functional outcome. Indeed, HIF-1α upregulation can lead to cell survival and thus may be a positive adaptive response in tendinopathy. The JNK/stress-activated protein kinases have been implicated in tenocyte apoptosis29 and while a moderate increase was noted in JNK/stress-activated protein kinases in our culture system this did not prove significant. At a mechanistic level we could not identify a functional role for ERK and p38 in the apoptotic response to hypoxia, which may suggest a mitochrondrial regulatory pathway as in synovial fibroblasts.49
There are limitations inherent in our study. First, age-related changes within the tendon samples could contribute to the degenerative picture and hypoxic cell expression seen in the matched subscapularis tendons. However, the lack of degenerative change on MRI and arthroscopic examinations suggests that the differences are truly at the cellular level, as suggested by our work. Second, the subscapularis tendon is functionally and organisationally distinct from supraspinatus and thus responds to mechanical loading in a different manner that may alter its cellular profile. Also control samples from subscapularis undergoing stabilisation may not be truly ‘normal’ controls, but are currently the best available control tendon sample and this is reflected by a Bonar score of 1. It is reassuring, however, that we found the same apoptotic and hypoxic cell markers in matched subscapularis tissue, indicating that the these responses may be uniform within joints subjected to tendon degeneration. In addition, having subscapularis samples from the same patient eliminates bias that may result from variation between individuals, and has previously been shown to be a useful method in sampling of tissues.50 Finally, the use of this in-vitro system while currently the ‘gold standard’ available to us may not accurately represent the in-vivo model and thus we acknowledge further animal work may be necessary.
On the basis of these results, we propose hypoxia as a critical pathophysiological regulator in tendon healing—a better understanding of its pathological cascade should lead to the development of cell-targeted treatment modalities for early supraspinatus tendinopathy.
References
Footnotes
-
Competing interests None.
-
Funding This work was funded by grants from the Arthritis Research UK Orthopaedic Clinical Research Fellowship and the Royal College of Surgeons of Edinburgh Cutner Fellowship.
-
Ethics approval This study was conducted with the approval of the South East Sydney Ethics Board, NHS Greater Glasgow and Clyde Research and Ethics Committee (West Section).
-
Provenance and peer review Not commissioned; externally peer reviewed.